视觉系统的normalization
视觉信号的神经通路
ref: <<神经科学:探索脑>>
- 视网膜 → 外侧膝状体核LGN →V1/初级视皮层*纹状皮层 →颞叶与顶叶
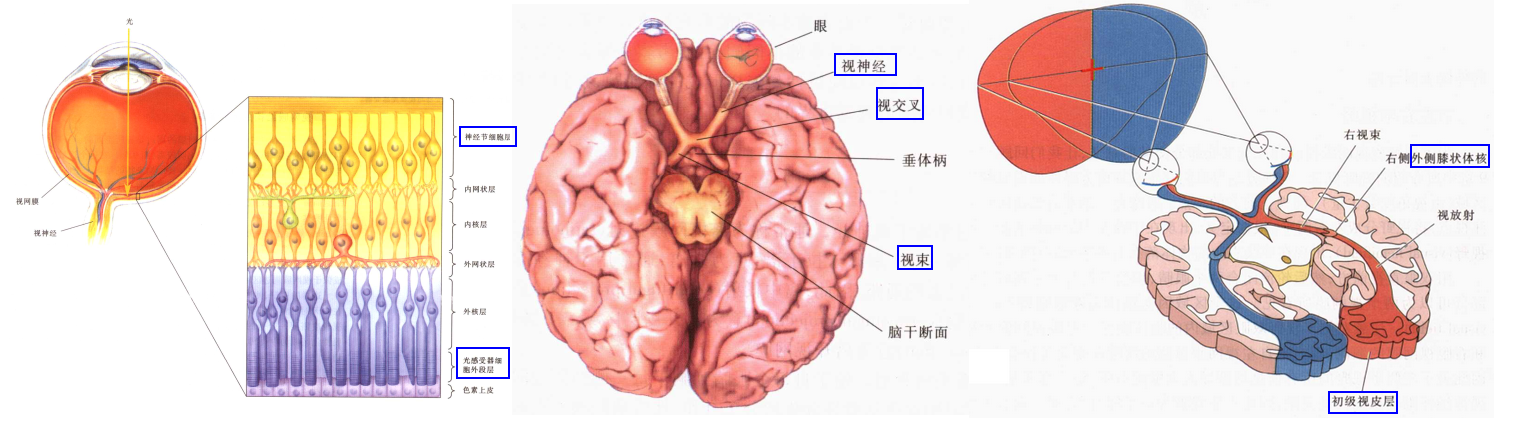
-
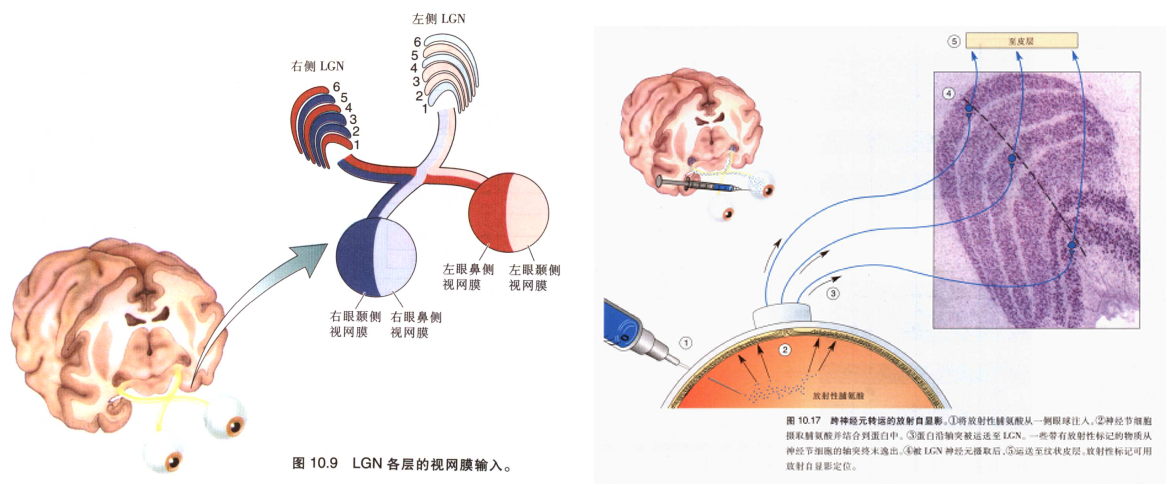
视觉内容在LGN的投射和实验证明
切片是垂直于皮层切的
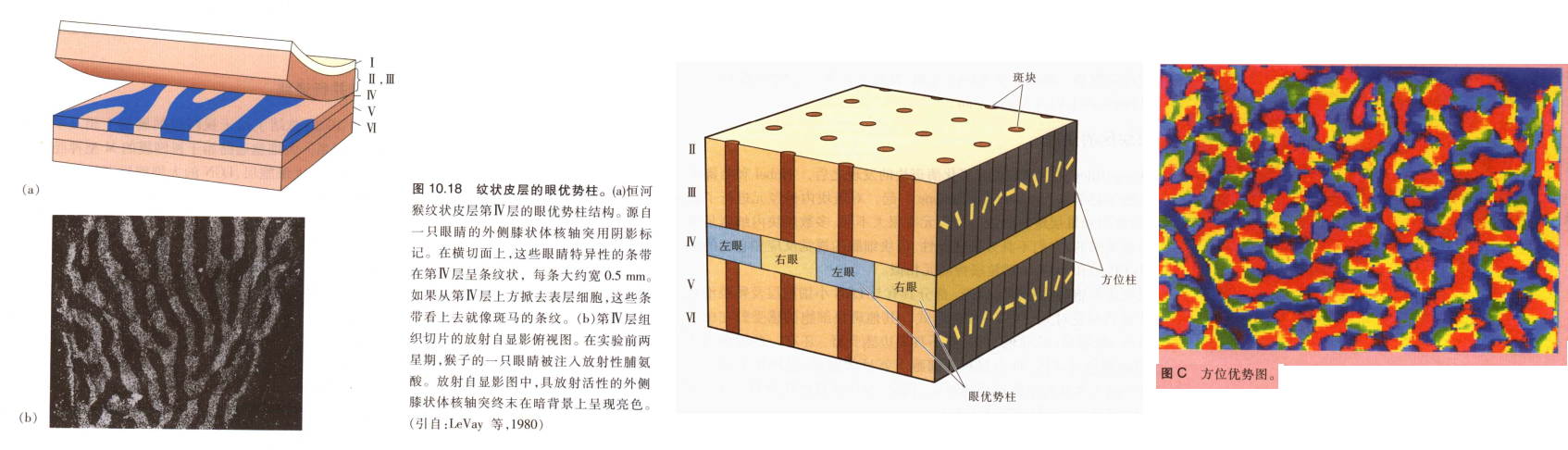
- 纹状体眼优势柱(左右眼) 和斑块区(方位)
在平行于皮层的切面方向,眼睛的轴突投射并非是均匀展开的,而是被分为一系列空间大小相等的条块,每个宽约0.5mm, 这些条块为眼优势祝 ocular dominance column 。
- 投影拓扑图
视觉通路的 normalization
https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1009028
Learning divisive normalization in primary visual cortex:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8211272/
Divisive normalization (DN) is a prominent computational building block重要的计算模块 in the brain that has been proposed as a canonical cortical operation. 大量的实验已经正式它在非线性神经元响应的重要性,DN也是处理自然刺激的重要组成部分
是一种在大脑中被广泛认为的基本计算方式。在这种机制中,单个神经元的发射率是其对输入的响应与其所在神经元池中所有神经元活动的总和的比值。这种模型被成功地用于描述从视觉和嗅觉系统的感觉处理到前运动和顶叶区域中与环境相关的价值编码等一系列任务。
例如,在视觉领域,除法归一化可以解释在主视觉皮层中,当视觉皮层中的神经元对刺激区域中的一个刺激做出反应时,如果周围区域有额外的刺激,该反应会被抑制。
人类的视觉皮层包括初级视皮层(V1,亦称 纹状皮层 (Striate cortex))以及纹外皮层 (Extrastriate cortex,例如V2,V3,V4,V5等)。在V1皮层中,有很多文献对其非线性响应特性进行了DN建模.
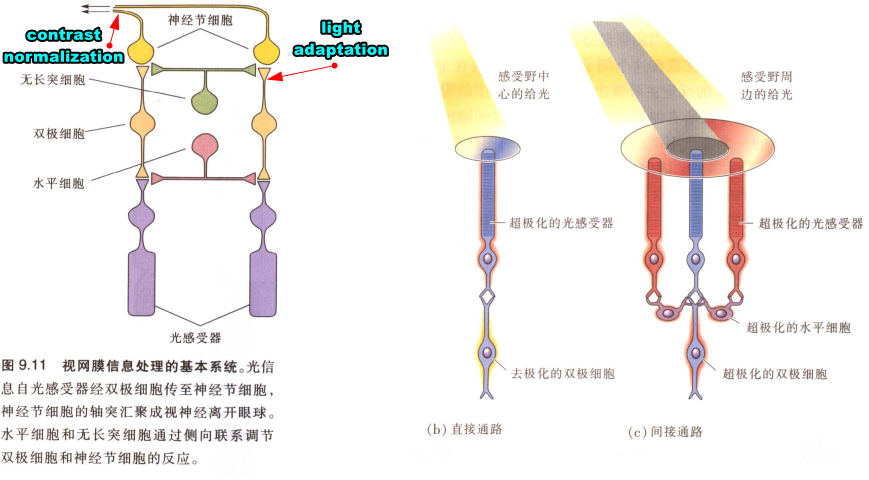
视网膜retina的标准化
Normalization as a canonical neural computation:https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3273486/
The retina needs to operate in a wide range of light intensities. Across visual environments (for example, an overcast night and a sunny day) light intensities range over 10 factors of 10(跨越了10个10倍的范围,10个数量级). Within a given visual scene the variation is typically smaller, but still more than a factor of 10 . 在平常的环境中,从白天到夜晚,亮度跨越了10个数量级,就算是在一个场景下,也有一个数量级.
The retina uses anatomical solutions (rod and cone photoreceptors and an adjustable pupil) for some of this wide range. 视网膜在解剖结构上通过调解rod 、cone、瞳孔适应这么广的范围.for the rest of the range, light adaptation adjusts the sensitivity of neurons through normalization在更广的范围中,则是通过normalization 调节神经元的敏感性
由此,在眼球上光适应的手段:
- rod,cone 的光感受器(非线性响应曲线)
- 瞳孔大小
- 神经元的normalization(局部区域和有限时间窗,即这种神经元的抑制效果需要在满足在短时间内同时发生,且是空间上邻近的神经元,可以认为是感受野抑制效应)
rod,cone 的光感受器(非线性响应曲线)
单个神经元非线性响应的曲线公式类似于这样:I是activity,R是response, 参数 γ/n/σ 控制曲线的形状,对于视觉来说 n一般=1
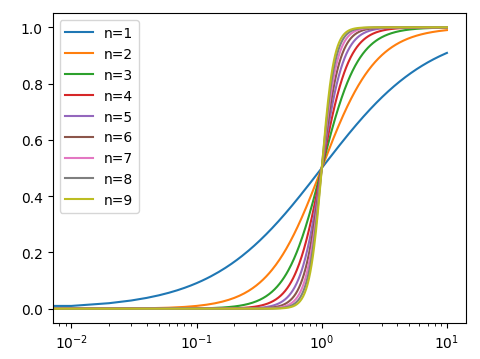
light adaptation
这里的Im表示 环境的整体背景强度,可通过求均值得到(the mean light intensity), 因此下标为m
这里的Im 虽然表示环境的整体亮度,但是在神经元层面可以理解为周边神经元的响应,由于环境变亮了,周围的神经元也整体变亮了,从而产生了抑制效应
看模拟的取下,当环境整体变亮时候,比如0.5倍率(变暗),1倍,1.5… 曲线右移动log(dx)
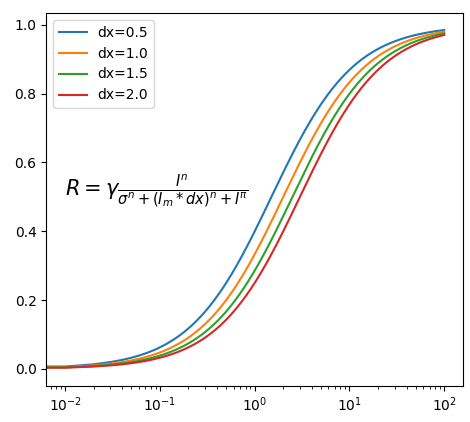
- 证明 曲线 近似右移lgdx:
原始响应曲线为:
亮度调整dx后曲线为:
当响应相同时R0=R1:
当 I0,I1 >> σn时:
得证
由此,我们可以解释,在不同光强下,看到的图片是很相似的. 比如下面两个栅格, 亮度成倍右移后,感受的直方图上分布是相似的
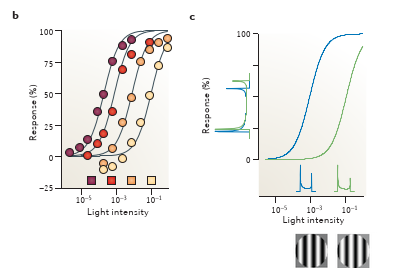
This adjustment of sensitivity approximates a neural measure of visual contrast 这种灵敏度调整策略使得在不同光强下,神经都可以测量对比度,即对比度恒常性 (Rm的 I 为Im下的响应),
contrast normalization
Conceptually, it is useful to consider contrast normalization as separate from light adaptation 29,30, but mechanistically the two stages may overlap in bipolar cells. 从概念上讲,将对比度归一化与光适应分开考虑是有用的,但从机制上讲,双极细胞可能在这两个阶段都有参与
Light adaptation is thought to operate in photoreceptors and bipolar cells, whereas contrast normalization is thought to occur in bipolar cells and ganglion cells. 光适应被认为在光感受器和双极细胞中起作用,而对比度标准化被认为发生在双极细胞和神经节细胞中
Under contrast normalization 在对比度标准化中, responses are no longer proportional to local contrast Cj (the output of the first normalization stage). Instead, the response Rj of neuron j is divided by a constant σ plus a measure of overall contrast:
Here, the weights wi (positive or negative) define the spatial profile of the summation field (typically, a centresurround difference of Gaussians), and the weights αk (positive) define the spatial profile of the suppressive field (typically, a large Gaussian) 通常 标准化的分母感受野更大一点
上述对比度标准化公式中,分布是标准差,这一关系式在LGN(lateral geniculate nucleus (LGN))中得到验证。(retina to LGN)
感受野大小的实验,是通过调节sinusoidal grating 的 size/window size/degree 大小验证的,
increasing grating contrast leads to response saturation when gratings are shown in a large window, but not when they are shown in a small window
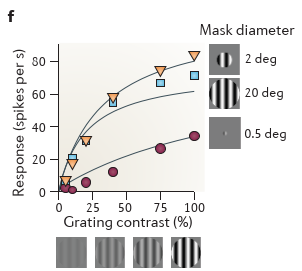
上述特性就是我们常说的感受野
神经节细胞类型:
- 按大小分
神经节细胞类型有大 小两种:
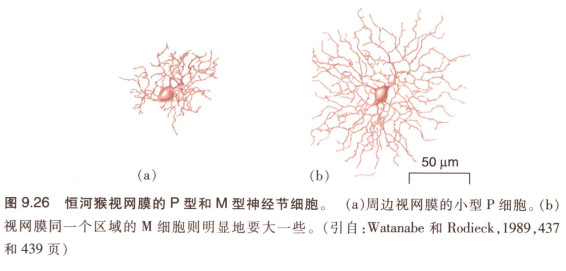
在恒河猴中 小的P细胞月占总神经节细胞90%,而M细胞月为5%,其余5% 为各种non M- non P的神经节细胞。
-
M细胞具有较大感受野,传到速率快,响应随刺激时间而减弱,对移动的刺激敏感→对瞬变敏感
-
P细胞具有较小感受野,和刺激时间同样长的持续放电,对细微之处敏感 →对细节关注
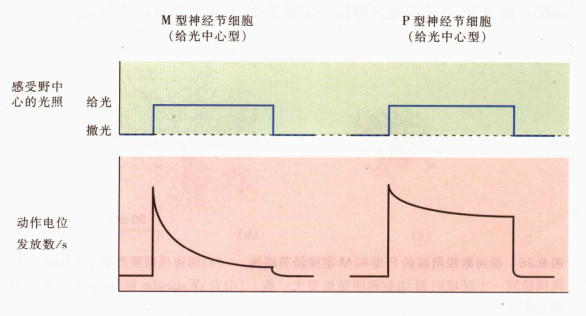
-
按功能分
颜色对立细胞 color opponent cell
- 红绿对立
- 黄蓝对立

V1视觉皮层/纹状体 primary visual cortex的标准化
从前面视觉信号通路的介绍中,我们可以了解到视觉皮层某个位置(大脑中某个三维坐标)的细胞,包含了视网膜成像中的位置(x,y)和方位角(θ)信息,这里的方位角是线条的角度。 这个细胞的响应分为: x 这里代表位置(x,y)
分子:用D(x,θ)代表 summation field 周边神经元的贡献
分母:用N(x,θ) 代表 suppressive field 周边神经元的抑制
具体公式表示为:
V1神经元 Here, we describe the normalization model for a population of V1 neurons differing in preference for stimulus position and orientation. 这种标准化因素使得能够响应更广泛的刺激。 指数n的区间通常是在 1.0 ~ 3.5
更一般化的公式:
但为了简化 ,会使用:
Normalization explains why responses of V1 neurons saturate with increasing stimulus contrast, irrespective of stimulus orientation, and therefore irrespective of firing rate. 归一化解释了为什么V1神经元的反应会随着刺激对比度的增加而饱和,而与刺激方向无关,因此也与放电速率无关(看不懂这句话,)
这里的γ可认为与优势方向有关,D是来自retina的contrast,由此公式又变为:
- θ代表优势方向:preferred orientation
- φ代表当前刺激方向: stimulus orientation
- c为contrast
- R 为response
因此对不同朝向, 他的响应只是 γ因子在调节, 对比度不受φ影响,γ不受c影响
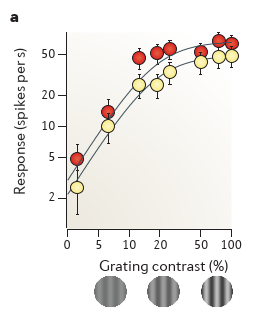
cross-orientation suppression
The responses of a V1 neuron to a test grating that drives responses are suppressed by superimposing on the test grating a mask grating that is ineffective in eliciting responses when presented alone — for example, because its orientation is orthogonal to the neuron’s preferred
orientation43,53–56. Normalization explains this effect because the suppression in the denominator increases with both test contrast ct and mask contrast cm, whereas the stimulus drive increases only with test contrast ct
The effect of increasing mask contrast, therefore, is to increase the denominator, which shifts the curve to the right on a logarithmic contrast axis. 右移的证明可以参考前面小节
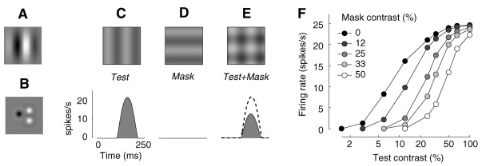
Transition from drive to suppression
Normalization also explains the more general case in which the mask does provide some drive to the neuron, for example, because its orientation is close to the preferred one (FIG. 3c). In this case, normalization makes an important prediction43,57: the mask should evoke activity when presented alone, but it should become suppressive in the presence of a more effective stimulus.
在单独只有次优方向(grating2)时,随着grating2 增长,神经元发放会增加,但比较弱
在单独只有最优方向(grating1),神经方法频率最高,但是随着grating2加入,反而会下降
grating 1 对比度越弱, 随着 grating2 下降越快
Grating 1 had optimal orientation and grating 2 had suboptimal orientation.
这一规律可从下列公式中推得:
w1 >> w2 measure the degree to which stimulus 1 and 2 drive the neuron (and for simplicity we are setting the exponent n to 1). When stimulus 1 is absent (c1 = 0), responses increase with stimulus 2 contrast. If instead stimulus 1 has sufficiently high contrast c1, one can ignore the term w2c2 in the numerator. Therefore c2 appears only in the denominator, where it exerts a purely suppressive effect.
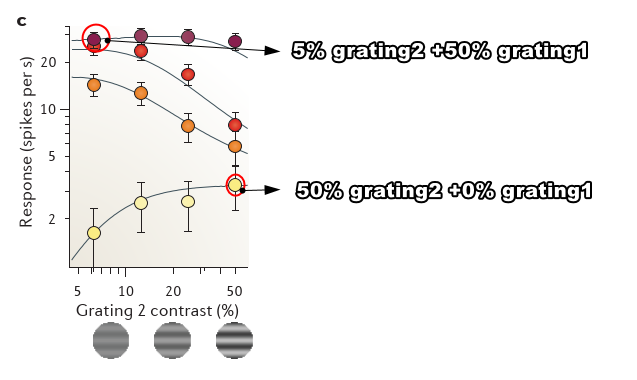
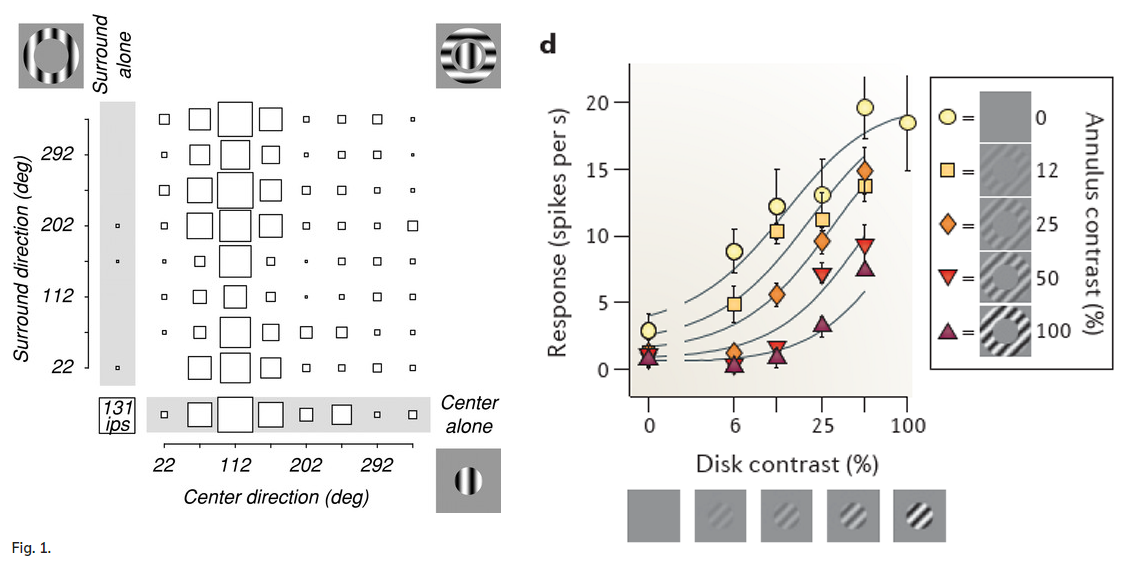
Surround suppression
原本对中心敏感的神经元会因为 周边 圆环(Annulus) 对比度的增加而受到抑制. 注意这里说的是神经元,而非人最终的感知.
https://journals.physiology.org/doi/full/10.1152/jn.00693.2001
summary
We have seen that divisive normalization is a widespread computation in disparate sensory systems, and that it may also play a part in cognitive systems (for example, those that encode value). Why is normalization so widespread? A tempting answer would be to see it as a natural outcome of a very common mechanism or network; a canonical neural circuit. However, there seem to be many circuits and mechanisms underlying normalization and they are not necessarily the same across species and systems. Consequently, we propose that the answer has to do with computation, not mechanism. Normalization is thought to bring multiple functional benefits to the computations that are performed by neural systems (BOX 2). Some of these benefits may be more important for some neural systems than for others.
Box2
- Maximizing sensitivity.
- Invariance with respect to some stimulus dimensions
- Decoding a distributed neural representation.
- Discriminating among stimuli
- Max-pooling (winner-take-all)
- Redundancy reduction.
除法归一化 divisively normalized
Learning divisive normalization in primary visual cortex: https://pubmed.ncbi.nlm.nih.gov/34097695/
-
x是刺激,
-
l 是某个神经元,输入信号yl 的 nl 次幂,而后经由 除法进行归一化,k∈K 是周围神经的权重
-
σl是semi-saturation constant, 以上所有参数非负
-
K是标准化神经元,pkl是weight
